
Зубов А. А. ЕСТЕСТВЕННАЯ ИСТОРИЯ ДРЕВНЕГО ЧЕЛОВЕЧЕСТВА
"Природа", 1998, № 1
|
|
|
Александр Александрович Зубов, доктор исторических наук, заведующий отделом антропологии Института этнологии и антропологии РАН. Научные интересы связаны с изучением систематики рода Homo. Монографии: "Этническая одонтология" (М., 1973); "Одонтология в современной антропологии" (в соавторстве с Н.И.Халдеевой. М., 1989); "Одонтология в антропофенетике" (в соавторстве с Н.И.Халдеевой. М., 1993)
ЭВОЛЮЦИЯ человеческого рода не протекала по одной прямой восходящей линии. Постоянный процесс видообразования порождал различные направления развития, возникали новые виды и подвиды, число которых сейчас составляет предмет острых дискуссий в антропологии. Ведутся они и по поводу разных моделей систематики гоминид.
Попробуем разобраться в естественной истории древнего человечества.
Антропологи иногда забывают, что систематика человека — это ветвь великого древа таксономии всего живого мира. Отчасти это связано с философскими установками, которые породили представление о непроходимой пропасти между гоминидами и остальными животными, о качественном отрыве человеческих существ от биологического субстрата в связи с возникновением новой модели жизни. По этой причине таксономические построения, относящиеся к гоминидам, оказались в значительной степени изолированными от зоологической систематики и ее основных принципов. В результате возникло положение, которое известный специалист по зоосистематике Г.Симпсон охарактеризовал как хаос в антропологической номенклатуре.
Каждый исследователь, обнаруживший какие-либо костные остатки ископаемых гоминид, стремился выделить новый вид или даже род, нередко преувеличивая таксономический ранг и эволюционную значимость находки. Бесчисленные "плезиантропы", "телантропы", "явантропы" и даже такие заведомо близкие формы, как питекантроп и синантроп представлялись таксонами родового ранга. Неандертальский человек фигурировал как отдельный вид.
В результате к 60-м годам нашего века было зарегистрировано 105 "новых" видов, из числа которых 33 сразу же были признаны неправомерными в соответствии с законами зоосистематики и номенклатуры, а 38 поставлены под сомнение как "неоправданные".
60-е годы принесли новые веяния во взгляды на классификацию палео-антропологических находок. Их изобилие побудило специалистов обратить больше внимания на повторяемость сходных черт, чем на бросавшиеся прежде в глаза индивидуальные различия. Медленно, но неуклонно дробление начало сменяться укрупнением, объединением и снижением таксономического ранга находок.
Это умонастроение захватило подавляющее большинство ученых и привело некоторых из них к изменению своих прежних теоретических позиций. Например, М.А.Гремяцкий, пересмотрев собственную более раннюю таксономическую схему, пришел к выводу о необходимости объединить в один род питекантропа и синантропа. Антропологи М. Ф. Нестурх, Г. Ф. Дебец, Э. Брайтингер, Э. Майр, Дж. Робинсон пошли еще дальше — объединили всех "истинных" гоминид в один род Homo, придав им ранг видов. Затем в семействе гоминид оказались и австралопитеки. Особенно радикальной была позиция Б.Кемпбелла: он выделил в пределах рода Homo только два вида — человека прямоходящего (H. erectus) и человека разумного (Н. sapiens), причем в последний включил и неандертальца, и человека современного физического типа как два подвида.
Многие антропологи (в частности и автор этой статьи) тогда восприняли идеи Кемпбелла как чрезмерный крен к "объединительству" — мол, извечный маятник качнулся в противоположную дроблению сторону. Однако известный антрополог Д. Джохансон предостерегает от чрезмерного укрупнения таксонов гоминид, мотивируя это недостаточной информативностью костных материалов, которые часто не позволяют адекватно оценить размах различий на уровне вида. Джохансон считает неандертальца отдельным видом и не включает в состав Н. sapiens. Словно в подтверждение его высказываний, появилась публикация английских специалистов, в которой они доказывают по митохондриальной ДНК, что неандертальцы и линия, ведущая к современному человеку, развивались независимо на протяжении более чем 500 тыс. лет.
Однако сейчас есть основания включить неандертальца в состав Н. sapiens в качестве подвида (H. s. neanderthalensis): многие факты пошатнули прежнее убеждение в низком интеллектуальном уровне и эволюционно примитивной организации этого гоминида. Оказалось, например, что скелет и осанка неандертальца из Ля Шапель-о-Сен (позвоночник, положение головы) фактически не отличались от осанки современных людей. Следовательно, его архаизм во многом преувеличивался.
Кроме того, в Сен-Сезер (Франция) костные остатки неандертальца найдены вместе с орудиями типа шательперрон (он свойствен верхнепалеолитическому человеку современного типа). Значит, резкие культурная и эволюционная границы между неандертальцами и современными людьми отсутствуют. А находка скелета неандертальского человека в гроте Кебара (Израиль), захороненного в могильной яме, как будто говорит даже о существовании какой-то похоронной обрядности и о довольно высоком уровне духовной жизни. Кстати, недавно обнаружено еще одно подтверждение высокого эволюционного уровня неандертальского человека: среди тех же останков из грота Кебара найдена подъязычная кость, фактически идентичная по строению такой же кости человека современного физического типа. Эта находка — существенный аргумент в пользу того, что человек из Кебара, бесспорно принадлежавший к неандертальской фазе эволюции, вероятно, владел членораздельной речью.
Взгляд на неандертальца как подвид Н. sapiens порождает ряд новых подходов к некоторым другим проблемам. Например, понятие "сапиентация" тогда приложимо уже к стадии формирования неандертальского человека, а начало этого процесса должно быть отнесено к среднему плейстоцену. Строго говоря, можно было бы пойти и дальше, распространив период сапиентации почти до истоков человеческого рода. Такие попытки делались. А. Тома еще в начале 60-х годов предлагал объединить всех гоминид, начиная с архантропов, т.е. питекантропов и синантропов, в один вид — Н. sapiens.
В самом деле, мы знаем, что архантропы еще 1.42 млн лет назад применяли огонь. По находке останков мальчика-архантропа из Нариокотоме-Ш (Кения) известно, что скелет Н. erectus и современного человека мало отличались ![]() .
.
Словом, объединительная тенденция нашего времени в теории антропогенеза — не просто дань всеобщему направлению мысли, не просто стремление восстановить равновесие и соответствие между систематикой гоминид и иерархией других зоологических таксонов, а объективная необходимость, диктуемая фактами. Очевидно, род человеческий — это непрерывно эволюционирующий таксон, который нелегко подразделить на сколько-нибудь обособленные "этажи" прогресса.
Мысль о возможности и вероятности метисации между представителями разных стадий эволюции наиболее определенно, пожалуй, впервые высказал тот же Тома. Он пришел к выводу, что морфологически промежуточные палестинские формы ископаемых гоминид могли быть не переходным звеном от неандертальца к современному человеку, а результатом метисации между ними. Это было смелое заключение, ведь в те времена еще не хватало фактических оснований для такого предположения. Гипотеза Тома не встретила всеобщей поддержки, тем более что подобная точка зрения в какой-то мере нарушала строгий порядок, установленный стадиальной теорией антропогенеза.
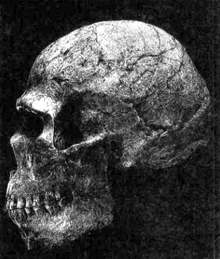 |
|
Череп раннего Homo sapiens из Палестины, сочетающий черты современного типа (намечающийся подбородоч-ный выступ, высокий свод) и неандертальского комплекса (довольно сильно развитое надбровье). |
Если для обоснования метисационной гипотезы в 50-х годах не было твердых доказательств, то теперь они есть. Факт сосуществования во времени, притом на близких территориях, представителей разных фаз эволюции гоми-нид уже не вызывает сомнений. Это доказывают дати-ровки костных остатков шанидарских неандертальцев (46 тыс. лет назад, Ирак) и черепов H. s. sapiens из грота Кафзех (92 тыс. лет назад, по другим данным даже 115 тыс. лет, Израиль). В Африке человек современного типа жил уже 120 тыс. лет назад, т.е. в период "пика" господства более архаичных форм. Особенно примеча-тельно, что примерно в то же самое время и на сравнительно близкой территории обитал очень примитивный представитель рода Homo, о котором мы знаем по находке черепа в Брокен-Хилл. Группа так называемых пренеандертальцев, к которым причисляют и человека из Брокен-Хилла, настолько архаична, что некоторые исследователи относят их к архантропам или предлагают включить эти формы в один вид H. heidelbergensis (название дано по первой находке близ Гейдельберга нижней челюсти одного из представителей этой группы). Следовательно, пренеандер-тальцы — столь примитивная ветвь человеческого рода — и люди современного физического типа сосуществовали в смежных регионах в течение тысячелетий.
Возможности для метисации были. В Африке, например, могли встретиться представители нескольких эволюционных стадий. Марокканские "питекантропы" дожили там до начала верхнего плейстоцена и были современниками неандертальцев, да и H.s.sapiens существовал уже несколько десятков тысячелетий.
А что происходило на территории Европы в тот период (36 тыс. — 25 тыс. лет до наших дней), когда древнее неандертальское население замещалось популяциями H. s. sapiens?
Судя по палеоантропологическим и археологическим данным, представители этого подвида пришли в Европу примерно 34 тыс. лет назад из Африки через Переднюю Азию. За несколько тысячелетий на сравнительно небольшой территории Западной Европы возникла весьма интересная ситуация: там сосуществовали "классические" неандертальцы, промежуточные формы и фактически сформировавшиеся популяции H. s. sapiens. Эти факты, по мнению разных исследователей, могут свидетельствовать либо о метисации, либо о трансформации неандертальских форм в современные.
Каковы же аргументы в пользу того или иного процесса?
Современный тип появляется как бы неожиданно и сосуществует с промежуточными и неандертальскими популяциями. Трудно предположить, чтобы трансформация протекала так стремительно. Конечно, можно возразить, что неандерталоидные черты исчезли не сразу, они сохранялись примерно до XXV тысячелетия, если не дольше (это подтверждают, например, находки на стоянке Сунгирь во Владимирской области). Однако даже 10 тыс. лет — период довольно незначительный для столь существенных морфологических изменений. Если допустить, что скорость трансформации была чрезвычайно высокой, то как объяснить наличие промежуточных черт (тот же Сунгирь) в течение долгого времени?
 |
|
| Реконструкция головы девочки из Сунгирского захоронения (работа Т. С. Сурниной |
Сторонники гипотезы трансформации приводят в качестве аргумента усиление черт сапиентного комплекса с течением времени. Но тогда получается, что человек современного типа возник в Европе независимо от других эволюционных ветвей, притом гораздо позже, чем в Африке. Я не придерживаюсь ортодоксальной позиции полицентризма, и мне трудно с этим согласиться. Словом, в Европе скорее можно представить картину метисации пришельцев с местным населением, нежели трансформацию неандертальцев в более прогрессивный таксон. Не выдерживает критики и гипотеза полного истребления неандертальского населения людьми современного типа: последние приобретают неандерталоидные черты уже позднее первоначальной миграции в Европу, т.е. примесь имеет местное происхождение.
Интересно, что большой миграционный взрыв, повсеместно захвативший популяции H. s. sapiens, приходится, видимо, на период 35—30 тыс. лет до наших дней. Именно к этому времени относятся находки костных остатков древних представителей "нашего" подвида вне Африки и Европы, включая даже островной мир (Япония, о. Окинава, 32 тыс. лет; о. Тайвань, 30 тыс. лет). Вероятно, тогда-то и закрепилось окончательно господство нового подвида в Старом Свете и он, вероятно, впервые вступил в пределы Нового Света. При таком огромном масштабе миграции могла происходить повсеместная метисация с локальными популяциями гоминид. Они и придали своеобразие потомкам разных волн мигрантов и способствовали формированию современной картины расовой дифференциации.
Но реализовалась ли метисация всюду, где для нее были предпосылки? Отнюдь. В Африке, например, первые популяции H. s. sapiens, очевидно, просто вытеснили более древних гоминид, почти не смешавшись с ними. Зато заселив Европу, пришельцы с востока скорее всего ассимилировали какую-то часть местного населения. В чем была причина столь разного поведения людей современного типа?
Можно допустить, что более 100 тыс. лет назад, когда H.s. sapiens столкнулся в Африке с поздними архантропами или представителями H. heidelbergensis, между этими двумя эволюционными стадиями все же существовал некий биологический (и, конечно, культурный) барьер, который препятствовал метисации. В Европе же барьер между прогрессивными восточными пришельцами и некоторыми популяциями неандертальцев был значительно менее выражен: культурный разрыв между ними, как думает большинство современных исследователей, не стал непреодолимым.
О метисации, происходившей в период активных миграций, можно судить по характерным особенностям современных рас, в частности по таким нейтральным и стабильным показателям, как одонтологические маркеры, т.е. особенности морфологии зубов. Например, лопатообразная форма резцов использована Ф. Вейденрейхом как доказательство прямой преемственности между синантропом и современными монголоидами. Я. Я. Рогинский как моноцентрист выдвинул альтернативную гипотезу. Он считал, что расселяющийся Н. sapiens приобрел некоторые черты (среди них и лопатообразные резцы) локальных популяций гоминид благодаря метисации. На востоке Азии такой формы резцы унаследованы от синантропа или его непосредственных потомков.
В принципе такая точка зрения оправдана. Но не следует забывать, например, что резцы европейских неандертальцев имели в основном лопатообразную форму и были очень сходны с резцами синантропа. Не унаследована ли эта форма зубов от неандертальца еще до широкого расселения человека современного типа на восток?
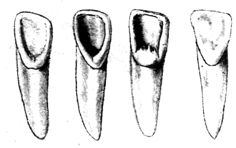 |
|
Типы верхних резцов человека (по: Ю. Мидзогути). Слева направо: типы 1, 2, 3 и промежуточный вариант |
У первых представителей человеческого рода — H. habilis — лопатообразная форма резцов отсутствовала, преобладал "западный" морфотип — как у современных европеоидов и негроидов Африки (по классификации японского одонтолога Ю. Мидзогути — это тип 1). Этот тип, видимо, существовал на африканском континенте в эпоху H. еrectus, включая поздние его варианты, в той или иной мере выражен у палестинских форм. В верхнем палеолите он очень четко представлен в остатках из Сунгирского погребения и, наконец, доминирует у современного населения всей западной части ойкумены (Европа, Африка). Создается впечатление, что в западных регионах чрезвычайно долго сохранялся африканский вариант структуры верхних резцов.
В Азии картина совершенно другая. Там, начиная с древнейших представителей H. erectus, господствуют лопатообразные резцы типа. Правда, эта форма обнаруживается и у европейских неандертальцев, характеризует некоторые палестинские варианты, а в западных регионах не сохраняется позже палеолита. В Азии же она гораздо сильнее влияла на формирование структуры резцов H. s. sapiens, но у современных монголоидов все же редко встречается в первоначальном классическом виде. Из этого следует, что западный и восточный одонтологические стволы фактически не развивались независимо, параллельно, если не считать, может быть, нескольких ограниченных во времени периодов изоляции. Вероятно, современный восточный расовый ствол формировался на основе западного за счет примеси местных азиатских форм с характерным для них вторым типом строения верхних резцов.
Но почему же считается, что метисация происходила в Азии, а не в Европе — с местными популяциями? Ведь краниологические материалы свидетельствуют о смешении H. s. sapiens с неандертальцами, причем известны даже ископаемые промежуточные формы. Кроме того, некоторые находки в Палестине имеют и одонтологические свидетельства метисации — средние варианты морфологии верхних резцов (синтез типов 1 и 2).
Вернее всего и в западном, и в восточном регионах ойкумены метисация происходила повсеместно и приводила к образованию таких "синтетических" морфологических вариантов. Но носители типа 1 на западе были многочисленнее, так как генетический вклад неандертальцев, а соответственно и морфотипа 2, намного меньше 50%: На востоке же более многочисленными были носители второго морфотипа, так что именно он получил перевес в последующих поколениях. В периоды относительной изоляции первичных надрасовых стволов образовавшиеся различия за счет генетико-авто-матических процессов привели к увеличению концентрации типа 1 на западе и типа 2 (в обновленном метисацией варианте) — на востоке.
Как мы видели, по распределению двух типов резцов можно заключить, что H. s. sapiens формировался при активном взаимодействии восточного и западного очагов эволюции гоминид. Происходило оно уже на ранних этапах формирования неандертальского человека, т.е., по современной терминологии, — на заре возникновения Н. sapiens. Следовательно, характер эволюции рода Homo на всех этапах его становления был "сетевидным" — с участием восточной и западной ветвей антропогенеза. Исключение, может быть, представляет временная изоляция этих ветвей в отдельные периоды существования H. erectus. Важно отметить, что "сеть" могла включать разные эволюционные "этажи", взаимодействовавшие между собой и вносившие свой генетический вклад в общий, единый генофонд рода Homo. Картину антропогенеза следует, таким образом, представлять как куст ветвей, соединенных между собой.
Ортодоксальные сторонники полицентризма представляют эволюцию рода Homo по-другому. По их данным, расогенетические линии достаточно четко маркируются признаками, которые показывают генетическую преемственность расовых типов, начиная с H. erectus. Не исключено, что в этой аргументации есть свой резон. В частности, отдельные проторасы, находившиеся на периферии древней ойкумены, могли без значительных влияний со стороны дать достаточно изолированные линии развития, приведшие к тому или иному своеобразному расовому типу, например линии австралийских аборигенов.
Такую форму эволюции в некоторых случаях, видимо, нельзя исключить, но она не везде была правилом. В частности, в Европе не могло быть подобных изолированных эволюционных линий с выраженной преемственностью форм на больших отрезках времени. В. В. Бунак проанализировал краниологический полиморфизм в Европе в период верхнего палеолита и пришел к выводу, что краниологические типы, соответствующие конкретным формам последующих эпох, здесь отсутствовали. "Расы" верхнего палеолита, по Бунаку, не были предковыми формами современных расовых типов, не имеют определенных ареалов и не могут считаться расами во всем объеме этого понятия. То была лишь часть эволюционной "сети", которая в конце концов привела к становлению более или менее стабильных форм нашего времени.
Подобный характер дифференциации краниологических типов подтвердил немецкий исследователь В. Хенке. Он изучил распределение 192 верхнепалеолитических и мезолитических черепов Европы по девяти измерительным признакам и обнаружил, что широкие и низкие черепа типа Кро-Маньон I и узко-высокие черепа типа Комб-Капель занимают противоположные полюсы. Совокупность проанализированных черепов в целом выглядела аморфной, в ней не проявлялась тенденция к образованию определенных комплексов. Не различались даже верхнепалеолитические черепа от мезолитических: диагностика эпохи дает лишь 57% правильных определений, т.е. невозможно уловить направленность изменений во времени. Мы застаем в данную эпоху в Европе картину морфологической пестроты, в основе которой лежит метисация, происходившая при достаточно большой по тому времени плотности населения.
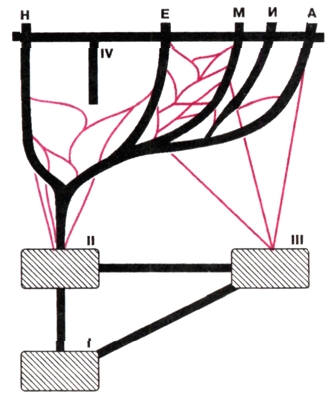 |
|
Схема формирования полиморфного внутривидового состава современного человечества. Н — негроиды Африки, Е — европеоиды, М — монголоиды, А — австра-лоиды, И — индейцы Америки. I — общеаф-риканский предок африканских (II) и азиат-ских (III) архантропов, IV — "перекрываю-щая" миграция Homo sapiens sapiens. Видно, что метисация (цветные линии) представите-лей разных эволюционных ветвей (черные линии), начавшись с архантропов, в дальней-шем усилилась, благодаря чему эволюцион-ное древо приобрело вид довольно густой сети. |
В проведенных статистических исследова-ниях анализировались только европейские мате-риалы. В Африке, не говоря уже об Австралии, наследование современным человеком более ранних, в частности верхнепалеолитических, форм выражено как будто определеннее. Так, черепа из Эльментейта, Фиш-хука, Флорисбада, относящиеся к верхнему палеолиту — мезолиту, имеют черты нынешних расовых типов Африки. Особенности современных вариантов монголо-идной расы, видимо, складывались по европей-скому типу, т.е. через густую "сеть" линий, в которой направления формирования этих вари-антов наметились сравнительно поздно. Об этом свидетельствуют два факта: нейтральный мор-фологический комплекс черепа из верхнего гро-та Чжоукоудянь, а также отсутствие сколько-нибудь выраженной совокупности монголоид-ных черт у южнокалифорнийской находки из Юхи 14 (21.5 тыс. лет). Впрочем, и все древнейшие краниологические материалы на территории Америки тоже их не имеют.
Если в Европе примерно в то же время был представлен нейтральный, мозаичный тип, который по краниологическим данным даже нельзя назвать европеоидным (пример — Сунгирь), то в Африке и Австралии гораздо раньше уже сложились черты будущих рас: экваториальный вариант, сходный с бушменским; прототасманийский комплекс; прототип грацильного австралоидного варианта.
Очевидно, что формирование современных краниологических комплексов шло по-разному и неодинаковыми темпами в каждом регионе. На большинстве территорий Евразии выделение расогенетических линий с непрерывной преемственностью сильно задерживалось из-за активной метисации при относительно большой плотности населения. В Африке, Юго-Восточной Азии (а затем и в Австралии) такие линии определились раньше и не подверглись значительному смешению. Напротив, древние местные формы, отчасти сложившиеся еще в период первичной дифференциации H. erectus, возможно, способствовали стабилизации линий. Метисация там могла быть узколокальной и не нарушала общих направлений расогенеза, а, наоборот, поддерживала их.
Человечество, вероятно, дважды делилось на восточную и западную части. Первый раз это произошло во времена H. erectus (или ранее); тогда и возникла "южная пара" очагов расообразования — в Африке и Юго-Восточной Азии. Во второй раз — в эпоху формирования H. s. sapiens в Африке, который расселился по всей ойкумене и смешался с местными популяциями; образовалась "северная пара" расогенетических очагов: первая — в Европе и Передней Азии; вторая — в Восточной и Центральной Азии, на территории Китая. Протоевропеоидный и протомонголоидный северые очаги активно взаимодействовали друг с другом и отчасти с древними южными центрами. Однако вблизи последних с давних времен формировались и локальные расовые типы. Они не испытали серьезных побочных влияний и сохранили в основных чертах местные направления расогенеза вплоть до наших дней.
В целом представленная картина генезиса современных рас опирается на активную метисацию и постоянное взаимодействие микроэволюционных линий ("сетевидная" эволюция), с той только оговоркой, что эти процессы не везде шли с одинаковой интенсивностью. Вследствие этого линии, ведущие к разным современным расовым типам, имеют неодинаковую древность и генетическую однородность.
Отстаиваемая мною позиция довольно существенно расходится с часто упоминаемой в популярной литературе гипотезой "митохондриальной Евы": якобы все современные расы произошли в Африке от одной прапопуляции около 200 тыс. лет назад, после чего не было никакой метисации с более примитивными гоминидами. Такой вывод противоречит данным одонтологии и краниологии, которые свидетельствуют, что в расовых типах современного человечества сохранился заметный отпечаток древнейшей дифференциации.
Однако между моей позицией и гипотезой "митохондриальной Евы" есть важное сходство: признается первостепенная роль "перекрывающей", сравнительно поздней африканской миграции древнего H. s. sapiens в формировании современного человечества.
Изложенная здесь точка зрения, будучи в общем моноцентристской (единый центр происхождения H. s. sapiens — Африка), сходится с полицентризмом в том, что локальные варианты древних гоминид сыграли ту или иную роль в генезисе основных расовых типов человечества. Каждая раса рассматривается как некоторый итог эволюции всего рода Homo в целом.
H. erectus признается большинством исследователей отдельным видом, однако поздние (H. heidelbergensis и некоторые неандертальцы), более прогрессивные формы, возможно, уже не были отделены биологическим барьером от Н .sapiens и могли внести свой существенный вклад в формирование рас современного человека. Расы, таким образом, имеют сложный генофонд, составленный разнородными и разноуровневыми микроэволюционными линиями. Они — продукт не только дифференциации, но и интеграции (метисации). При этом расы, видимо, различаются по "густоте" сети взаимодействующих линий, явившихся итогом локальных направлений микроэволюции.
Вероятно, европеоиды и монголоиды — это наиболее "смешанные" расы: в их генофонд влилось множество микроэволюционных линий, включая неандерталоидные и даже более ранние эволюционные формы, генетическое взаимодействие которых было весьма интенсивным и длительным. Значит, эти две большие расы должны считаться более "молодыми" по сравнению с негроидами и особенно австралоидами, в формировании которых метисация сыграла меньшую роль. Не следует забывать при этом, что нынешние расы "вдвойне" родственны друг другу: ведь для всего рода Homo корень един — африканский, а миграции на разных эволюционных уровнях смешали генофонды разделившихся ранее ветвей.
Особое место в расогенетической схеме занимают американские индейцы. В прежних классификациях их нередко просто относили к большой монголоидной расе. Однако подробное и всестороннее исследование не подтверждает эту точку зрения. Аборигенные группы Америки пришли в Новый Свет через участок суши в Беринговоморье еще до того, как сформировались черты современных монголоидов и характеризуются нейтральным типом. А он произошел от недифференцированного единого восточного ствола, давшего впоследствии монголоидов и австралоидов. Современных американских индейцев следует отнести к особой "американоидной" расе, их нельзя включить ни в одну из ныне существующих больших рас. Разнообразие типов аборигенного населения Америки — это следствие нескольких миграций со все большим удельным весом монголоидного компонента в каждой следующей волне. Соответственно самая древняя из них имела наиболее австралоидный облик.
Японский генетик М. Ней считает, что подвид H. s. sapiens впервые разделился на две микроэволюционные ветви примерно 100 тыс. лет назад, причем одна из них была представлена протонегроидами, а другая — недифференцированным европеоидно-монголоидным стволом, разделившимся значительно позже, около 60 тыс. лет назад.
Одновременно Ней отмечает большую генетическую близость субрасовых таксонов как внутри европеоидной, так и монголоидной рас. Его результаты хорошо подтверждают уже высказанную точку зрения об интенсивном генном обмене между большими расами Евразии, основные современные черты которых стали окончательно складываться позже верхнего палеолита. Другие же расовые стволы уже давно эволюционировали — формировались свойственные им в настоящее время морфологические комплексы.
Выводы Нея, казалось бы, противоречат идее деления человечества на восточный и западный стволы. Однако никакого противоречия не будет, если допустить возможность интенсивной метисации представителей евро-монголоидной ветви с более ранними гоминидами как на западе, так и на востоке.
"Сетевидный" характер эволюции на всех стадиях антропогенеза, на первый взгляд, не укладывается в принятую сейчас модель систематики рода Homo, в которой архантропы выделются в самостоятельный вид — H. erectus. Но, во-первых, не все соглашаются с этим: в настоящее время африканских архантропов объединяют в особый таксон — человек трудящийся H. ergaster), а азиатских относят к H. erectus. Во-вторых, между антропологами нет согласия и по поводу верхней границы существования H. erectus (ergaster). Одни проводят ее примерно на уровне 200 тыс. лет до наших дней, включают в этот таксон промежуточные формы H.heidelbergensis и называют их "прогрессивными архантропами". Другие же предпочитают относить этих пренеандертальцев к отдельному виду. Вследствие таких раз-ночтений время, когда архантропы сошли с "эволюционной сцены", оценивается по-разному: 700 тыс. лет назад, 400 тыс. и, наконец, 200 тыс. (возникновение архантропов все согласно относят к 1.6 млн лет назад). Как бы то ни было, таксон уровня архантропа существовал по меньшей мере около 1 млн лет, а скорее всего — дольше, особенно в Азии.
Разумеется, за этот огромный период он должен был претерпеть существенные изменения как за счет сапиентации, так и благодаря образованию боковых ветвей (некоторые из них оказались тупиковыми). Можно думать, что из огромного числа весьма разнообразных по морфотипу и эволюционному уровню популяций архантропов далеко не все они внесли генетический вклад в эволюционный потенциал формирующегося вида Н. sapiens. Архаичные тупиковые группы, ставшие его современниками, остались за чертой репродуктивной изоляции. Их можно причислить к виду H. erectus вместе с ранними, примитивными формами этого таксона. Более продвинутые в эволюционном отношении потомки архантропов образовали вид H. heidelbergensis (некоторые исследователи называют его ранним архаичным сапиенсом).
Сейчас невозможно решить, где проходит видовая граница, разделяющая эту группу, видимо, чисто морфологический критерий здесь недостаточен. Упомянутая трактовка таксономической структуры H. erectus позволяет примирить идею Кемпбелла, выделившего архантропов в отдельный вид, с моделью, в которой признается возможность метисации некоторой части представителей этой систематической группы с более прогрессивными формами. Вряд ли можно согласиться с тотальным исключением восточной или западной ветвей архантропов из числа возможных предков Н. sapiens. Мозаичный одонтологический тип европейских неандертальцев скорее свидетельствует о наличии в генофонде последних как "западных", так и "восточных" черт, унаследованных от разных групп H. erectus.
Итак, каково же мнение исследователей в отношении систематики рода Homo?
Приведенные здесь рассуждения дают аргумент в пользу сохранения трехвидовой структуры рода Homo: H. erectus, H. habilis и Н. sapiens. К первому виду, наверное, следует отнести ранние, а также пережиточные формы архантропов, которые скорее всего были репродуктивно несовместимы с Н. sapiens. Однако сомнительно, чтобы в отдельные виды были выделены африканские архантропы и неандертальцы. Особняком стоит также Н. heidelbergensis.
Но таксономический статус вида H. habilis, окончательно не решен: некоторые исследователи разделяют его на H. habilis и H. rudolfensis. Тогда число видов в пределах рода Homo возрастает до семи: H. habilis, H. rudolfensis, H. erectus, H . ergaster, H. heidelbergensis, H. neanderthalensis, H. sapiens.
Если принять эту таксономическую модель, то придется признать, что почти на всех стадиях эволюции рода Homo происходило ветвление с образованием новых видов и тупиковых форм.
Несомненно, в цепи промежуточных вариантов, соединяющих австралопитека и H. erectus, должны были существовать формы, подобные H. habilis, но трудно сказать, составляли ли они реальный вид. Если в новых исследованиях, в частности связанных с определением древности вида H. erectus, будет доказано, что он существовал около 2 млн лет назад, то формы, относимые сейчас к H. habilis, окажутся либо современниками H. erectus, либо даже "моложе" его, а следовательно, образуют боковую ветвь эволюции. Человек умелый при этом остался бы, вероятно, в роду Homo, но появились бы новые, непредвиденные проблемы в теории антропогенеза и систематике гоминид.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект 96-06-80159).