|
|
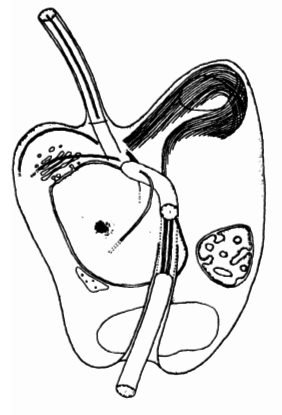
| Корешковые микротрубочки Bicoso-eca maris образуют широкую ленту, поддерживающую область клеточного рта. Из Moestrup, 1982. | |
Микротрубочки - жесткие арматурные прутья, которые в комплексе с белком диенином способны к очень быстрым движениям. Они имеются у всех известных эукариот. У некоторых групп простейших, например, Эвгленозоа, микротрубочки могут образовывать жесткий наружный каркас, проходящий под пелликулой или входящий в состав пелликулы. Если клетка имеет вырост или впадину, то они, как правило, поддерживаются микротрубочками. Во многих случаях микротрубочки определяют положение ядра и органелл в клетке ![]() , расхождение ядер во время клеточного деления. Однако бывают организмы, которые в основных стадиях своего жизненного цикла не имеют микротрубочек. Микротрубочки состоят из шариков (глобул) белка тубулина, шарики могут полимеризоваться с образованием нитей, которые соединяются с образованием трубочек
, расхождение ядер во время клеточного деления. Однако бывают организмы, которые в основных стадиях своего жизненного цикла не имеют микротрубочек. Микротрубочки состоят из шариков (глобул) белка тубулина, шарики могут полимеризоваться с образованием нитей, которые соединяются с образованием трубочек ![]() . Итак, обладая шариками тубулина, клетка эукариот может быстро собрать и столь же быстро разобрать жесткие конструкции.
. Итак, обладая шариками тубулина, клетка эукариот может быстро собрать и столь же быстро разобрать жесткие конструкции.
Как определяется место сборки? В разных местах клетки могут находиться ЦОМТы - центры организации микротрубочек. Примером ЦОМТов являются центриоли - во время деления ядра они инициируют появление "нитей митотического веретена" - микротрубочек, прикрепляющихся к хромосомам и разводящих их к разным полюсам клетки. Центриоли - остаток или производное от базального тельца жгутика. ЦОМТы не возникают ниоткуда - при делении клетки дочерние базальные тельца "отпочковываются" от материнских. Обычно клетки, особенно примитивные, обладают ограниченным набором ЦОМТов, который во многих случаях является важным систематическим признаком. Например, простейшие, ранее рассматривавшиеся в составе класса Солнечников, оказались двумя совершенно неродственными группировками. Обе имеют аксоподии - длинные узкие выросты цитоплазмы, постоянство формы которых поддерживается плотными тяжами микротрубочек - аксонемами. У актинофридных солнечников ЦОМТы расположены на ядерной мембране, а у центрохелидных солнечников в середине клетки находится центропласт (центросома, аксопласт), от которого и отходят аксонемы. Центропласт сходной структуры (характерная трехслойная дисковидная пластинка, от которой разбегаются микротрубочки) обнаруживается у Слизистых грибов и лобозных амеб, что позволяет говорить о их возможном родстве.
 |
|
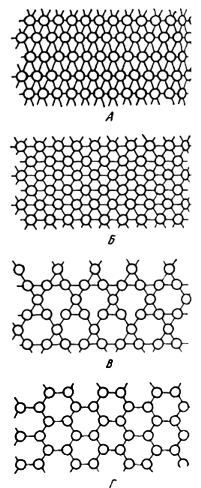
Различные формы сое-динения микротрубочек |
Микротрубочки, в свою очередь, способны полимери-зоваться с образованием крупных, иногда заметных в световой микроскоп образований - например, аксостилей. У разных простейших трубочки могут соединяться с двумя и более соседними трубочками, поэтому говорят о двухвалентных, трех- и более валентных микротрубочках. Валентность трубочек - признак отряда или класса. Многовалентные трубочки радиолярий и солнечников создают жесткие неветвящиеся аксонемы. Наоборот, микротрубочки мягких сетчатых ретикулоподий фораменифер расположены рыхло и не образуют линейных связей.
Если трубочки соединены друг с другом посредством белка динеина, они могут скользить друг вдоль друга и развивать значительное усилие. Подобное скольжение обеспечивает биение жгутиков и изгибание аксостиля и аксоенемы. В клетках животных микротрубочки могут обеспечивать транспорт веществ или мелких частичек. Быстрые движения микротрубочки осуществляют только в жгутике, где они организованы в уникальную 9+2 структуру. Считается, что жгутик возник в ходе эволюции только однажды и все жгутиконосные клетки произошли от одного предка.
Микрофиламинты также способны к быстрым движениям, но не образуют жестких структур. Они обеспечивают токи цитоплазмы в клетках и амебоидные движения. Микрофиламенты обычно образованы актином и миозином - белками, способными скользить друг относительно друга. Они есть у всех эукариот. У животных они обуславливают сокращение мышц. Сложные упорядоченные актомиозиновые комплексы имеют характерную поперечную исчерченность, которая хорошо заметна в поперечнополосатых мышцах. Актомиозиновые волокна - мионемы - обычны у инфузорий, способных иногда с поразительной быстротой сокращать отдельные части клетки. В растительных клетках они, самостоятельно или во взаимодействии с микротрубочками, обеспечивают движение хлоропластов или токи в цитоплазме. В последнем случае микрофиламенты прикрепляются к мембранам эндоплазматической сети и, смещая их, "раскручивают" все содержимое клетки. Сходный ток тонкого слоя цитоплазмы вдоль аксонем солнечников и радиолярий обусловлен взаимодействием микротрубочек и микрофиламентов, когда микрофиламенты движутся по микротрубочкам, как по рельсам. Движение амеб становится невозможным при блокаде действия акто-миозинового комплекса.
Цитоскелет обеспечивает и более направленное движение вещества в клетке. Например, основное назначение апарата Гольджи - образование пузырьков с секреторным веществом; в некоторых случаях это материал для постройки наружной оболочки или домика. В нем, например, синтезируются чешуйки и волоски, покрывающие снаружи многие клетки. Как они выводятся наружу? Пузырьки связываются с микрофиламентами внутреннего скелета, котрые выносят их в нужное место на наружной мембране.
Подобно пучкам микротрубочек, микрофиламенты способны образовывать более или менее рыхлые комплексы, которые также выполняют скелетную функцию. Микрофиламенты - более тонкие образования, чем микротрубочки. Они хуже выявляются электронномикроскопическими методами, поэтому менее изучены. Но там, где их исследовали подробно (в основном это многоклеточные организмы) их роль в организации формы клеток исключительно велика.
Помимо формы клеток, микрофиламенты определяют также расположение поверхностных белков. Известная метафора - поверхностные белки напоминают айсберги, плавающие в море липидной мембраны - неверна. Большинство поверхностных белков занимают на мембране строго определенное место, потому что связаны микрофиламентами с внутреними скелетными образованиями клетки. Те участки клеточной поверхности, которые способны “прилипать” к субстрату, укреплены микрофиламентами. С их помощью клетка может, “подтягиваясь”, перемещаться. Пищевые частицы прикрепляются к белкам-рецепорам, расположенным в мембране, что приводит к активации связанной с рецепторами системы микрофиламентов и втягиванию участка мембраны внутрь клетки. Так осуществляется фагоцитоз и пиноцитоз - заглатывание клеткой твёрдых частиц или пузырьков жидкости.
В некоторых случаях подвижные микрофиламенты образованы другими, плохо изученными белками. Поскольку эукариоты, в отличие от прокариот, практически неспособны к биохимической эволюции, развитая система движения, альтернативного "актино-миозиновому", имеет большое система-тическое значение.