


В. Н. Сергеев, Э. X. Нолл, Г. А. Заварзин ПЕРВЫЕ ТРИ МИЛЛИАРДА ЛЕТ ЖИЗНИ: ОТ ПРОКАРИОТ К ЭВКАРИОТАМ "Природа", 1996, № 6 |
|
|
|
|
||
| Владимир Николаевич Сергеев, кандидат геолого-минералогичес-ких наук, старший научный сотруд-ник Геологического института РАН. Область научных интересов — остатки микроорганизмов докем-брия и нижнего кембрия и воз-можности их биостратиграфичес-кого использования. | Эндрю Херберт Нолл, про-фессор Гарвардского универси-тета, действительный член Аме-риканской академии наук. Автор более 200 работ по остаткам древнейших микроорганизмов, изотопии пород и стратиграфии докембрийских отложений. | Георгий Александрович Заварзин, доктор биологичес-ких наук, профессор, заведую-щий отделом микробных сооб-ществ Института микробиологии РАН. Область научных интере-сов — глобальная микробио-логия. |
В Солнечной системе Земля - планета биологическая, отличающаяся сложной и разнообразной жизнью, которая сформировалась за долгую историю. Как свидетельствует палеонтология, эволюция, начавшаяся по меньшей мере 3,5 млрд. лет назад, шла с меняющейсяскоростью, когда относительно короткие периоды драматических перемен разделялись длинными этапами застоя. В привычной палеонтологической летописи последних 545 млн. лет изменения были особенно быстрыми. В этом возрастном интервале на границах между пермью и триасом, между мелом и палеогеном вымерло множество растений, животных, микропланктона, а выжившие получили возможность развиваться по новым путям. Во времена так называемого "кембрийского взрыва" большинство групп беспозвоночных увеличило свое разнообразие в океане, а многие группы и виды научились строить минерализованные скелеты. Глубоко и бесповоротно изменились экосистемы и биогеохимические циклы.
Естественно, что эти эволюционные события привлекают внимание биологов и палеонтологов; однако они охватывают только последние 15% длинной истории жизни. Более столетия назад Чарльз Дарвин писал в "Происхождении видов": "На вопрос, почему мы не находим богатых ископаемых, принадлежащих <...> периодам докембрийской системы, я не могу дать удовлетворительного ответа <...>. Явление остается необъясненным в настоящее время и может быть использовано как аргумент против приводимых здесь взглядов".
Только за последние 40 лет накопились новые данные в биологии, геологии, палеонтологии, позволившие существенно дополнить докембрийскую летопись Земли. Геологи большее внимание стали уделять древнейшим осадочным породам Земли, а появление изотопных методов датировки позволило реально оценить временные масштабы докембрийской истории. Глубокое понимание отличий природы растений и животных от бактерий и одноклеточных эвкариот привело к выводу, что ископаемые в докембрийских породах, вероятнее всего, должны быть представлены микроорганизмами. Поэтому палеонтологи, прекратили поиски все более древних трилобитов и с успехом стали искать остатки водорослей и бактерий. Во многих докембрийских сланцах были обнаружены сплющенные остатки микроорганизмов, в кремнях — тоже микроорганизмы, но сохранившиеся в трехмерном объеме, а в позднедо-кембрийских породах — многоклеточные водоросли и простые бесскелетные животные.
Задача нашей статьи — вкратце обсудить основные периоды биологической эволюции на докембрийской Земле и показать, как в течение первых трех миллиардов лет формировалась современная биосфера, а примерно 1 млрд. лет назад возникло основное разнообразие эвкариотных организмов. Наш тезис состоит в том, что палеонтологическая летопись может быть дополнена сравнительным экологическим подходом, взятым из микробиологии, а также данными молекулярной биологии, что позволяет получить более ясную картину событий далекого прошлого.
Следует добавить, что в настоящее время существуют международная и российская стратиграфические шкапы периодизации основных событий в докембрии (см. рис.1). В нашей статье мы постарались не обидеть ни одну из них.
Согласно молекулярной филогении, которая изучает эволюционные отношения между живыми организмами на основании нуклеотидных последовательностей, первые организмы были прокариотами — археями и бактериями. Ранняя жизнь базировалась на окислительно-восстановительных реакциях веществ, поступающих из глубины Земли в гидротермальных источниках. Восстановителем служил Н2, может быть СО, окислителем — СО2, S0; продуктами реакции соответственно были — СН4 и H2S. Такие экосистемы, которые используют поступающие из недр Земли неорганические вещества, не имели решающей роли в определении лика планеты: в геологической летописи нет следов эпох, характеризующихся сероводородной или метановой атмосферой.
Со временем, однако, некоторые микроорганизмы научились потреблять продукты обмена других организмов. Так сформировались микробные сообщества с замкнутым круговоротом веществ, для распространения которых важным фактором стало развитие фотоавтотрофии. Это был принципиальный этап в создании биосферы и ее эволюции в целом.
На современной Земле есть некоторые местообитания, где нет растений и животных, но процветают бактерии. Проблема состоит в том, чтобы оценить насколько такие области могут служить моделью древнейшей биосферы. Особое внимание привлекают три местообитания: глубоководные гидротермы, термальные источники и гиперсоленые лагуны. Все они интенсивно изучаются в поисках аналогий с прошлым, а развивающиеся в них сообщества рассматриваются как реликтовые, которые выжили несмотря на изменения окружающей среды и появление высших организмов.
Пожалуй, лучшим аналогом ранних экосистем Земли можно считать глубоководные океанические выходы термальных вод, если, конечно, исключить круговорот, приносящий воды современного океана с кислородом и частицами органического вещества. В термальных выходах обитают гипертермофильные микробы, среди которых имеются формы, принадлежащие по молекулярным признакам к древнейшим филогенетическим ветвям древа жизни. Однако мы вправе задать вопрос: могли ли они создать функционирующую биосферу, пригодную для дальнейшего развития жизни?
В теплых термальных источниках, где доминирующий газ — углекислота, развиваются богатые микробные сообщества, которые можно рассматривать как "природные эксперименты", моделирующие взаимодействие микробов с дегазирующей Землей. Здесь процветают сообщества прокариот с циано-бактериями как первичными продуцентами и полной цепью термофильных организмов, обеспечивающих круговорот вещества в системе: автотрофные термофильные организмы ассимилируют СО2 и начинают круговорот углерода в экосистеме; с ними сопряжен обмен других организмов, разлагающих органическое вещество. Все это говорит о том, что термофильное микробное сообщество способно образовать корень древа биоразнообразия и создать плацдарм жизни, преобразившей геосферу в биосферу. В архейских и раннепротерозойских осадочных и вулканогенно-осадочных толщах были обнаружены гидротермальные источники, но до сих пор никто не пытался найти там остатки термофильных сообществ или следов их метаболической активности.
С другой стороны, палеонтологи установили, что появление на Земле уже 3.5 млрд. лет назад фотосинтеза дало возможность жизни колонизовать участки поверхности, далеко удаленные от гидротермальных выходов. В Австралии и Южной Африке породы этого возраста содержат микрофоссилии, строматолиты (органогенно-осадочные структуры, образованные микробным сообществами) и геохимические свиде тельства присутствия сложных микробных сообществ с автотрофными, скорее всего фотоавтотрофными, первичными продуцентами. Сегодня аналогичные сообщества можно наблюдать в соленых лагунах, где развиваются галофильные цианобактериальные маты. Эти строго структу-рированные сообщества отличаются широким разнообразием микроорганизмов; продуцентами опять-таки служат цианобактерии, со здающие в мате плотный покров хлорофилла, который перехватывает фотосинтетически активную радиацию.
У нас нет никаких оснований предполагать, что бактериальные обитатели реликтовых сообществ сколько-нибудь серьезно изменились со времени своего возникновения. Согласно геоло гическим и биологическим данным ранние экосистемы были главным образом прокариотными и анаэробными Тем не менее геологическая летопись жизни в течение примерно первого миллиарда лет пока еще недостаточно документирована, чтобы осветить начало эволюции, когда анаэробные прокариоты приступили к созданию современных экосистем на Земле.
Если жизнь впервые появилась в условиях низкого содержания кислорода, то когда же древнейшая атмосфера и океаны стали пополняться кислородом и какие Последствия это имело для экосистем? Ответ на эти вопросы дают палеонтологическая и геологическая летописи палеопротерозоя (карелия), изученного гораздо лучше, чем архея.
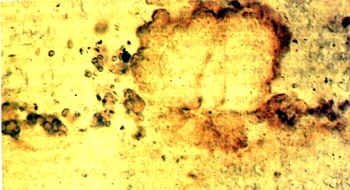 |
Рис.2. Коккоидная цианобактерия Eoentophysalis belcherensis Hoflnann из отложений мезопротеро-зойской котуйканской свиты Анабарского поднятия Сибири.Возраст ~1.4 млрд. лет. Здесь и далее фото авторов. | |
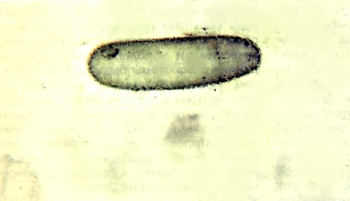 |
Рис.3. Эллипсовидная микрофоссилия рода Archaeo-ellipsoides Horodyski et Donaldson — фоссилизированная акинета (репродуктивная спора) ностоковых цианобактерий из отложений мезопротерозойской котуйканской свиты Анабарского поднятия Сибири. Возраст ~1.4 млрд. лет. |
Позднеархейские и ранне-палео-протерозойские кремни содержат микрофоссилии, которые, возможно, являются остатками цианобактерии. Однако в чуть более молодых породах (около 2.2—2 млрд. лет назад) обнаружены уже бесспорные цианобактерии. Примечательно, что эти первые ископаемые цианобактерии почти неотличимы от современных видов, живущих в сходных физических условиях. Например, Eoentophysalis belcherensis (рис.2), известная из многих палео- и мезопротерозойских отложений приливной зоны (tidal flats), очень похожа на современную цианобактерию Entophysalis major. В тех же древнейших отложениях приливной зоны находят микрофоссилии, относимые к роду Archaeoellipsoides (рис.3), которые представляют собой сохранившиеся репродуктивные споры (акинеты) цианобактерий, весьма сходных с акинетами современного рода Anabaena. Открытие акинет в палеопротерозойских отложениях интересно в свете современных данных молекулярной биологии. Согласно молекулярной филогении, устанавливаемой по последовательности нуклеотидов в 16S рибосомальных РНК (рРНК), образующие акинеты цианобактерий составляют самую верхнюю ветвь эволюционного древа цианобактерий. Таким образом, обнаруженные акинеты указывают, что основное разнообразие цианобактерий было достигнуто ко времени, когда мы впервые находим их бесспорные ископаемые остатки. Широкое распространение цианобактериальных сообществ в палеопротерозое подтверждают строматолиты, которых становится значительно больше на шельфе континентальных платформ 2.5—2 млрд. лет назад.
Экспансия цианобактерий оказалась решающей в развитии атмосферы и гидросферы Земли потому, что только эти организмы способны использовать воду для фотосинтеза. В цианобактериальном фотосинтезе молекулярный кислород образуется как побочный продукт, и его накопление в атмосфере и океанах преобразовало биосферу. Конечно, простая продукция кислорода еще не могла привести к атмосферной революции, поскольку большая часть образованного кислорода могла уйти в ходе биологических и химических процессов окисления. По имеющимся данным, Земля в палео-протерозое претерпела крупные тектонические изменения: образовались большие стабильные платформы, в мелководных шельфовых областях которых стало возможно долгосрочное захоронение органического вещества. Высокая скорость погребения органического вещества в палеопротерозое (о чем говорит аномально высокое изотопное соотношение углерода в органическом веществе и карбонатах), видимо, совпала с уменьшившимся потоком восстановленных вулканических газов. В результате этих событий 2.2—2 млрд. лет назад уровень атмосферного кислорода повысился с 1% до 10—15% от современного уровня. Такому резкому изменению в составе атмосферы соответствуют и изменения в древних корах выветривания с распространением минералов, образующихся при окислении кислородом.
Переход к кислородной атмосфере совпадает с почти полным исчезновением железистых кварцитов с магнетитом и появлением красноцветных пород. Это один из наиболее грубых, но наглядных геологических индикаторов наличия свободного кислорода и окислительных условий. Одновременно происходит перестройка и в цикле серы: в палеопротерозое сульфатредукция становится доминирующей, о чем свидетельствует изменение изотопного состава основных соединений серы в отложениях этого возраста. Поскольку циклы серы и железа несовместимы из-за образования сульфидов железа, то окисление сульфида аэробными бактериями в сульфаты моря стало возможным только около 2 млрд. лет назад после прекращения отложения железистых кварцитов.
Итак, в палеопротерозое биосфера Земли значительно изменилась: возникли новые, богатые кислородом экологические ниши на суше и в мелководных прибрежных участках, в то время как бескислородные экосистемы продолжали процветать в глубоких анаэробных частях бассейнов. Два миллиарда лет назад сформировалось полное функциональное разнообразие бактерий, необходимое для поддержания биогеохимических циклов на Земле в форме, близкой к современной. Начался следующий этап развития жизни.
Данные о последовательности нуклеотидов в 16S рРНК позволяют предположить, что эвкариоты как одна из трех главных ветвей эволюционного древа жизни имеют историю почти столь же длительную, как прокариоты. Однако древнейшие эвкариоты могли быть простыми анаэробными гетеро-трофами, которые не имели значения в ранних экосистемах. Эвкариоты смогли достичь своего эволюционного развития только в результате эндосимбиоза с бактериями. Одна из групп ранних эвкариот включила протеобактерию, способную к аэробному дыханию. Со временем эти "гости" стали биологически интегрированными с их хозяевами, создав композитную форму жизни. Потомки этих бактерий до сих пор существуют как митохондрии в клетках всех современных организмов, в том числе и наших с вами. Другие ранние эвкариоты дополнительно захватили цианобактерии, которые сходным путем превратились в хлоропласты и дали эвкариотным водорослям возможность фотосинтезировать.
Поскольку необходимым предварительным условием развития аэробных бактерий было распространение богатых кислородом экосистем, можно полагать, что искать ископаемых эвкариот нужно в породах более поздних, чем геохимические индикаторы, свидетельствующие о наличии кислородной атмосферы. Однако на практике идентификация древнейших остатков эвкариотных микроорганизмов в палеопротерозойских отложениях сталкивается с множеством трудностей. Дело в том, что низшие морфологически простые одноклеточные эвкариоты внешне почти неотличимы от прокариот и распознаются в основном по присутствию митохондрий и хлоропластов. К сожалению, эти органеллы не сохраняются в ископаемом состоянии, поскольку после гибели клетки ее протопласт сжимается до крошечных размеров, который у фоссилизированных микрофоссилий часто присутствует как темное небольшое шаровидное включение. (Интересно, что на начальном этапе изучения докембрийских микрофоссилий большинство подобных включений принималось за клеточные ядра, а сами микроорганизмы — за эвкариоты.)
Важным критерием, позволяющим различить про- и эвкариоты, служит размер, поскольку максимальный диаметр прокариотных клеток составляет около 60 мкм. Однако в ископаемом состоянии за остатки крупных клеток можно ошибочно принять слизистые оболочки, окружающие колонии некоторых цианобактерий, которые достигают в диаметре иногда нескольких сантиметров. К несомненно эвкариотным микроорганизмам относятся микрофоссилии сложной морфологии, прежде всего формы с шипами и выростами, нехарактерными для прокариот. Но остатки подобных микроорганизмов встречаются только вблизи границы мезо- и неопротерозоя, видимо, значительно позже появления первых эвкариот. Древнейшие микрофоссилии, с определенной долей вероятности относимые к эвкариотам, находят в породах с возрастом 1.7—1.9 млрд. лет. Они представляют собой большие, до нескольких сантиметров в диаметре, сплющенные тела сферических организмов, возможно, эвкариотного происхождения. И хотя эти отпечатки могут оказаться остатками не отдельных крупных клеток, а пустыми оболочками, окружавшими колонии прокариот, наличие эвкариот в сообществах палеопротерозоя подтверждается присутствием в битумах этого возраста молекул биомаркеров (стеранов), характерных только для некоторых представителей ядерных организмов.
Итак, хотя взятые отдельно доказательства слабы и допускают альтернативные интерпретации, в сумме они свидетельствуют, что эвкариоты, по-видимому, были широко распространены 17—1.9 млрд. лет назад. Биологическая принадлежность этих древнейших ядерных организмов в целом неопределенна ввиду простой морфологии, не связывающей эти ископаемые с конкретными ныне живущими группами протист. Вполне вероятно, что некоторые из них могли принадлежать к вымершим группам ранних эвкариот.
Вслед за важными биологическими и геологическими изменениями в палеопротерозое наступила длительная, сравнительно спокойная эра ме-зопротерозоя (1600—1000 млн. лет). Ассоциации микрофоссилии из окрем-ненных карбонатов приливно-отливной зоны, где преобладают цианобактерии, сходны как с палеопротерозойскими комплексами, так и с сообществами современных микроорганизмов. В сланцах этого возраста обнаружены многочисленные проблематичные микрофоссилии, — возможно, остатки эвкариотного фитопланктона. Помимо них на трех континентах найдены спиральные макроскопические отпечатки, которые почти наверняка принадлежат эвкариотам. Однако более разнообразные и сложно построенные остатки эвкариот в мезопротерозойских отложениях отсутствуют. По малопонятным причинам Земля почти не менялась в течение длительного времени, и смена биологических форм и увеличение их разнообразия были ограничены. Но в кажущихся простыми и статичными клетках эвкариот происходили молекулярные и ультраструктурные изменения, которые привели к эволюционному взрыву в конце эры.
Принципиальным биологическим событием в течение мезонеопротеро-зойского перехода (средний—поздний рифей) стало бурное увеличение разнообразия "высших" эвкариот и появление новых форм. Это изменение не было мгновенным: оно началось в позднем мезопротерозое и продолжалось, по-видимому, почти всю неопротерозойскую эру. Причины этого взрыва пока неясны, хотя, возможно, он связан с эволюцией цитоскелета и возникновением классических (митотических) форм полового размножения. В результате в течение короткого времени возникли почти все основные группы водорослей и простейших, а также предки животных, грибов, растений, что резко изменило природу и сложность биотических сообществ.
Древнейшие остатки эвкариотных организмов, которые, безусловно, отно-сятся к ныне существующим основным группам, представлены красными банги-евыми водорослями (рис. 4 ![]() ), сохранившимися в окремненных карбонатах приливной зоны из позднемезопротерозойской или ранненеопротерозойской формации Хантинг (Северная Америка).
), сохранившимися в окремненных карбонатах приливной зоны из позднемезопротерозойской или ранненеопротерозойской формации Хантинг (Северная Америка).
Далее в стратиграфической последовательности следуют находки зеленых (рис. 5 ![]() ) и, возможно, хромофитовых водорослей. Тогда же морфологически сложные микрофоссилии, обычно обозначаемые как акритархи (рис. 6,7
) и, возможно, хромофитовых водорослей. Тогда же морфологически сложные микрофоссилии, обычно обозначаемые как акритархи (рис. 6,7 ![]() ), начали быстро эволюционировать. Точное таксономическое положение акритарх неясно, потому что морфологически они весьма близки к разным линиям протист. Возможно, некоторые из них ныне вымерли, а другие вполне могут оказаться сохранившимися репродуктивными цистами празинофитных зеленых водорослей. В породах неопротерозоя появляются остатки и других протист, например — похожие на вазы микрофоссилии рода Melanocyrillium (рис. 8
), начали быстро эволюционировать. Точное таксономическое положение акритарх неясно, потому что морфологически они весьма близки к разным линиям протист. Возможно, некоторые из них ныне вымерли, а другие вполне могут оказаться сохранившимися репродуктивными цистами празинофитных зеленых водорослей. В породах неопротерозоя появляются остатки и других протист, например — похожие на вазы микрофоссилии рода Melanocyrillium (рис. 8 ![]() ), а также одноклеточные организмы Eosaccharomyces, напоминающие мицелий некоторых низших грибов. Более убедительные находки грибов относятся к отложениям верхнего неопротерозоя — венда, где они встречаются совместно с обильными остатками многоклеточных животных и бессосудистых растений.
), а также одноклеточные организмы Eosaccharomyces, напоминающие мицелий некоторых низших грибов. Более убедительные находки грибов относятся к отложениям верхнего неопротерозоя — венда, где они встречаются совместно с обильными остатками многоклеточных животных и бессосудистых растений.
Палеонтологические свидетельства о взрыве разнообразия эвкариот поддерживаются независимыми данными палеобиохимии и молекулярной биологии. Многочисленные находки биомаркеров говорят о возможном присутствии значительного количества зеленых, красных и хромофитовых водорослей в неопротерозойских экосистемах. Новейшие данные по молекулярной филогении ныне существующих протист также указывают, что появление разнообразных высших эвкариот произошло относительно поздно в истории группы. Этот большой взрыв ("big bang") эвкариотной эволюции дал предков трех ветвей многоклеточных организмов, доминирующих в современной биосфере: животных, растений, грибов (рис. 9 ![]() ). Таким образом, данные палеонтологии, палеобиогеохимии и молекулярной биологии согласуются между собой, предполагая, что основное веерообразное развитие эвкариот началось около 1 млрд. лет назад, в неопротерозое (рис.10).
). Таким образом, данные палеонтологии, палеобиогеохимии и молекулярной биологии согласуются между собой, предполагая, что основное веерообразное развитие эвкариот началось около 1 млрд. лет назад, в неопротерозое (рис.10).
Палеонтологическая летопись документирует появление эвкариот с определенной морфологией и сохраняемыми органическими частями. Однако многие ныне живущие мелкие и простые эвкариотные клетки почти не сохраняются в осадках. Некоторые из них, например инфузории, образуют определенные биомаркеры, наличие которых в породах может говорить об их присутствие и без структурно сохранившихся остатков. Но для многих простых эвкариот сведения о биохимическом составе их компонентов недостаточны, и поэтому в биогеохимических исследованиях конкретная цель поиска, остается неясной. Очевидно, видимая экспансия эвкариот в неопротерозое составляла только вершину айсберга, и более широкая экспансия протист не отмечена в геологической летописи.
Косвенные данные дают представление о возрастающем значении эвкариот в экосистемах, но и они не увеличивают наши знания о таксономическом разнообразии древних эвкариот. Например, образующие акинеты цианобактерии, которые обычны в отложениях мезопротерозоя, вероятно, жили в береговых водоемах с пониженной соленостью. Но их находки в отложениях неопротерозоя редки, несмотря на то, что эта группа широко распространена в современных водоемах. (Anabaena и некоторые другие формирующие споры водоросли хорошо знакомы, видимо, почти всем читателям, поскольку они — один из основных виновников "цветения воды".) Однако сейчас в прибрежных мелководных условиях, в которых обнаружено большинство остатков акинет мезопротерозоя, доминируют эвкариотные водоросли. Значит, редкость находок ископаемых акинет в отложениях приливных отмелей возрастом моложе 1000 млн. лет косвенно указывает на распространение более конкурентоспособных эвкариот, вытеснивших отсюда цианобактерий. Сходным образом, наблюдаемое сокращение строматолитов в разрезах неопротерозоя, вероятно, также говорит об экспансии морских эвкариотных водорослей и, позднее, животных.
В целом прямые и косвенные ископаемые свидетельства говорят о том, что хотя эвкариоты как группа возникли рано в истории Земли, распространились и стали доминирующими компонентами экосистем только около границы мезопротерозоя и неопротерозоя. Этим они создали предпосылку для следующей революции морских животных в начале фанерозоя и к переходу в конечном итоге к биосфере современного типа.
Наши представления о древнейшей жизни, основанные на ископаемой летописи, безусловно неполны. Только немногие фрагменты ее просматриваются с некоторой ясностью, но даже и достаточно хорошо сохранившиеся ископаемые отражают состояние жизни главным образом в ограниченных береговых областях, где и сегодня господствуют цианобактериальные маты; глубоководные моря и поверхность суши пока остаются белым пятном. То, что нам открывается, это не полный ландшафт докембрия, а скорее колодец в прошлое, и картину ранней палеонтологической летописи можно сравнить с наблюдением за освещенной верандой. Мы видим, как проходит жизнь на веранде, но можем только догадываться о том, что происходит внутри дома и на улице. К счастью, входящие и выходящие люди могут сказать наблюдателю очень многое относительно окружающей жизни.
Далее, мы можем расположить наши наблюдения в соответствии с общей схемой филогенетической информации, которая поступает из сравнительных биологических и молекулярных исследований живых организмов. Кроме того, современные цианобактериальные маты лагун, термальные источники, содовые озера служат местообитанием реликтовых микробных сообществ, которые дают важную таксономическую и экологическую информацию о возможных условиях жизни в отдаленном прошлом.
Таким образом, наиболее примечательная особенность палеонтологии докембрия состоит не в том, что наше понимание так неполно, но в том, что мы так много узнали за прошедшие 40 лет. Несмотря на неполноту летописи, мы знаем, что прокариоты создали сложные экосистемы уже 3.5 млрд. лет назад; мы знаем, что накопление кислорода в атмосфере в течение палеопротерозоя способствовало новому взрыву биоразнообразия, включая возникновение путем симбиогенеза эв-кариотной клетки, способной к аэробному дыханию; мы знаем, что эвкариоты быстро распространились около 1000 млн. лет назад; мы знаем, что за последующим подъемом содержания атмосферного кислорода появились крупные животные и настала современная эра. Эта картина показывает, что жизнь эволюционировала вместе со средой обитания на Земле, "биологическая" и "физическая" Земли объединены в одну систему, в которой обе стороны участвуют в биогеохимических циклах. Например — в цикле углерода: благодаря парниковому эффекту в атмосфере летучие компоненты воздействуют на климат; благодаря образованию карбонатных осадков он сопрягается с циклами других элементов и определяет среду обитания для морских организмов; благодаря захоронению органического вещества и его окислению цикл углерода определяет содержание кислорода в атмосфере. Когда события в развитии углеродного цикла привели к повышенному содержанию кислорода, жизнь эволюционировала с образованием все новых и разнообразных форм. Но они не вытеснили более ранние организмы; скорее они добавились к ним, ведя к большей сложности экосистем, которые продолжают до сих пор основываться на фундаменте бактериальных процессов, возникших в начале истории планеты.
Работа выполнена при поддержке гранта РФФИ 95-05-14575.