Медников Б.
Москва, "Советская Россия", 1975
|
|
Наряду с мутационным процессом второй фактор эволюции по Четверикову — колебания численности популяций, которые он образно называл «волнами жизни».
Каждый из нас наблюдал «волны жизни» воочию. Ранней весной редко можно увидеть случайную перезимовавшую между оконными рамами муху, а в августе мы страдаем от их назойливости. Год на год не приходится — в одно лето нет житья от комаров, а в другое они редки. Вспышки численности некоторых видов имеют воистину планетарный характер. Таковы, например, массовые миграции саранчи в годы ее усиленного размножения, годы «мышиной напасти», путешествия многочисленных стад полярных пеструшек — леммингов.
Некоторые из этих вспышек цикличны: они имеют сезонный, годовой или многолетний период (часто, хотя порой и без достаточного обоснования связываемый с солнечной активностью). Другие же апериодичны. Это вспышки численности организмов, попавших в новую среду обитания без достаточного количества врагов, эпидемии гриппа и иных заболеваний, с быстротой пожара распространяющиеся по планете нашествия вредителей из других стран.
Колорадский жук у себя на родине, в Северной Америке, никогда не достигал высокой численности, питаясь дикими растениями из семейства пасленовых. Печальную славу злостного вредителя он завоевал на европейских картофельных полях. То же можно сказать и об американской виноградной тле — филлоксере. Континент Евразии не остался, впрочем, перед Америкой в долгу, «наградив» ее непарным шелкопрядом, воробьями и длинным списком сорняков. Вспомним также нашествие кроликов и кактусов опунций на Австралию.
Отнюдь не всегда такие подъемы численности видов вредоносны. Бурное развитие в Каспийском море акклиматизированного там азовского червя — нереиса значительно увеличило кормовую базу рыб. Это в свою очередь компенсировало оскудение Каспия, наступившее в результате зарегулирования стока Волги. Не будь нереиса, осолонение Северного Каспия нанесло бы каспийским рыбам непоправимый ущерб. Никто не может возразить и против «нашествия» на нашу страну американской мускусной крысы — ондатры, расселившейся от Камчатки до Мурмана, от северной Сибири до Средней Азии (правда, по некоторым данным, она вытесняет ценного зверя — выхухоль). И все же испортить неразумным вселением сложившуюся структуру сообществ растений и животных гораздо легче, чем улучшить ее. Нелишне напомнить, что акклиматизации нереиса в Каспии предшествовала многолетняя работа. Сейчас с развитием авиации, облегчающей перевозки, порой случается и такое, что сначала акклиматизируют, а потом скромно уступают друг другу честь этого замысла. Дело доходит до курьезов, когда вместе с ценными объектами (а то и вместо них) завозят совсем нежелательные. Так, вместе с ценными растительноядными рыбами в Среднюю Азию завезли головастиков чернопятнистой лягушки с Дальнего Востока (уж этого-то можно было избежать!).
Много можно было бы рассказать интересного и о причинах таких колебаний численности, когда в период максимума она больше, чем в период минимума в миллионы раз. Обычные океанские волны никогда не достигают такой высоты, как «волны жизни»! Некоторые из них имеют характер автоколебательного процесса, как в генераторах радиоволн. Так, в период вспышки численности мышевидных грызунов прежде изолированные маленькие популяции сливаются в одну большую, возрастает вероятность контактов. Это приводит к быстрому распространению болезнетворных бактерий (например, чумы и туляремии), массовым эпизоотиям и резкому спаду численности. Гребень волны сменяется провалом, популяция колеблется, как маятник.
Однако сейчас нас интересует эволюционное значение таких колебаний. В первую очередь «волны жизни» резко расшатывают генофонд популяции, изменяя численность всех аллелей, разрушая сложившееся харди-вейнберговское отношение их. А это приводит в конце концов к изменению свойств популяции — так расшатывание выпадающего зуба облегчает процесс избавления от него. При спаде «волны жизни» численность одних мутаций возрастает, а значит, возрастает вероятность встречи рецессивов в гомозиготах; скрытые аллели проявляются в фенотипах и подпадают под действие естественного отбора. Новая «волна жизни» будет состоять из особей с измененным генофондом.
Другие мутации при спаде волны могут просто выпасть из генофонда. Этот эффект образно называют «горлышко бутылки» (или эффект бутылочного горлышка) — далеко не все аллели могут пройти к новым поколениям через минимум численности. Например, если при спаде численности лисиц из популяции исчезнут особи — носители генов черно-бурой окраски, чернобурок в этой популяции уже не будет.
Не менее важно и то обстоятельство, что при подъеме «волны жизни» прежде изолированные мелкие популяции объединяются в единую, начинается интенсивный обмен генами. Ареал такой популяции расширяется, периферийные части попадают в новые условия, в которых они еще не обитали. Эти enfants perdu (потерянные дети — так во французской армии называли сторожевое охранение на передовой) часто становятся предками новых форм, подвидов и видов (разумеется, если им повезет в жизненной борьбе). При спаде волны они нередко оказываются отрезанными от основного ареала и продолжают свое существование с тем генофондом, который у них имеется. А он, как правило, неадекватен общему генофонду всей популяции. Следующий подъем численности восстанавливает прерванную связь, но соединившиеся популяции порой уже «не узнают» друг друга, они уже генетически изолированы.
|
|
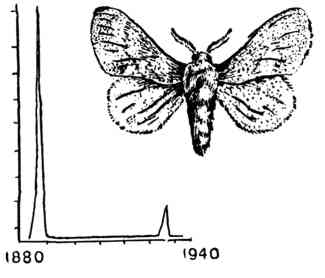
| Пример «воли жизни». Колебания численности зимующих гусениц соснового коконопряда за 60 лет. |
Наиболее четко это проявляется в феномене, описанном Четвериковым, который много позже эволюционист-теоретик Э. Майр назвал «принципом основателя». Нередко новый ареал (например, отдаленный остров) заселяют случайно занесенные туда несколько единичных особей нового вида. Так, вся западноевропейская популяция ондатры ведет начало от всего лишь пяти особей, выпущенных в начале века вблизи Праги. Пределом начала популяции, по-видимому, может быть одна оплодотворенная самка. Естественно, она одна, и даже несколько особей, не может хранить весь генофонд родительской популяции. Эволюция этого вида на новом месте должна пойти по-другому. Наряду с изменившимися условиями существования «принцип основателя» приводит к формированию островных рас, подвидов и видов.
В последнее время группа японских исследователей детально изучала популяции крыс на многих островах Океании. Эти грызуны — подлинный бич романтических атоллов — успели с момента вселения пройти большое расстояние по пути дивергенции (расщепления). Некоторые оказались уже генетически изолированными.
Однако колебания численности популяций сами по себе еще не могут привести к видообразованию, к становлению приспособленных к внешней среде форм. Они изменяют в популяции относительную численность аллелей; это несомненно, но несомненно и то, что эти изменения ненаправленны.
Тимофеев-Ресовский Н. В., Воронцов Н.Н., Яблоков А. В.
Москва, "Наука", 1977
|
|
Следующий элементарный эволюционный фактор, который может быть назван «популяционные волны», имеет совершенно иную природу, нежели мутационный процесс; в то же время, подобно мутационному процессу, он по природе своей статистичен.
Еще в 1905 г. С. С. Четвериков опубликовал чрезвычайно интересную работу под заглавием «Волны жизни». В этой работе им было показано, что у всех живых организмов все популяции всегда подвержены количественной флуктуации численности входящих в них особей. Характер, причины и размах таких колебаний численности могут быть очень различными; существенно лишь то, что в данном случае идет речь о флуктуациях, т. е. колебаниях в положительную и отрицательную стороны, сменяющих одна другую более или менее регулярно, а пе о постоянных, векторизованных процессах. Причины таких флуктуации могут быть весьма различны. В одних случаях они связаны с сезонной периодикой (например, у многих насекомых и других животных с относительно коротким жизненным циклом, а также у многих однолетних растений), в других — с более длинными и менее регулярными периодами, связанными с флуктуациями климатических условий или урожая кормов. Наконец, в ряде случаев могут иметь место апериодические, основанные на случайных «катастрофах», изменения численности, возникающие в результате наводнений, лесных пожаров, исключительных засух или морозов и т. д. Такие катастрофы ведут к резкому сокращению численности, затем восстанавливающейся и переходящей на уровень нормальных флуктуации.
Теперь можем с полной определенностью утверждать, что в биосфере Земли не существует популяций, не подверженных количественным флуктуациям, хотя диапазон этих колебаний может быть очень различен. У некоторых организмов (например, у многих древесных пород) такие колебания в связи с большой длительностью отдельных поколений протекают чрезвычайно медленно, часто ускользая от нашего непосредственного наблюдения.
У других организмов они выражены весьма ясно и бросаются в глаза каждому; достаточно вспомнить о колоссальных сезонных колебаниях численности многих животных. Попытки количественных, правда, в большинстве случаев еще не очень точных оценок приводят к отношениям численности особей в соответствующих популяциях в сезоны пика и спада до порядков 1 : 1000 000, а иногда и более. Такие колоссальные колебания характерны, однако, лишь для популяций организмов с короткими жизненными циклами и соответственно несколькими поколениями за вега-тационный сезон.
Почти те же пределы размаха колебаний численности в периоды пика и спада наблюдаются во многих случаях несезонных, периодических или непериодических колебаний численности многих «вредителей» или видов, связанных со значительными колебаниями кормовых объектов, в свою очередь зависящих иногда от сложных констелляций многих факторов; широко известны массовые инвазии насекомых-вредителей (непарного шелкопряда, вредной черепашки, бабочки-монашенки, гессенской мухи, саранчи и многих др.), общеизвестны годы «мышиной напасти», известны резкие «волны жизни», связанные с урожаем и неурожаем кормов у белок, зайцев, леммингов, известны связанные с резкими волнами жизни других животных колебания численности хищников и паразитов (у лис, песцов, хищных птиц, многих насекомых, клещей и т. д.), известны колебания численности некоторых однолетних растений. Наконец, столь же общеизвестны случаи массовых вспышек численности видов, попадающих в новые регионы, в которых отсутствуют их естественные враги. Такие апериодические резкие вспышки постепенно, по мере «освоения» местными биогеоценозами новых пришельцев, переходят в обычные, естественные флуктуации численности (кролики в Австралии или в Новой Зеландии, канадская элодея в Палеарк-тике, ондатра в Европе, американские сельскохозяйственные вредители в Евразии, домовые воробьи в Америке и т. д.). Менее известны и изучены аналогичные резкие апериодические «волны жизни», возникающие в результате «катастроф». Обычно в результате уничтожения одних видов резко возрастает численность других, ранее подавляемых или ограниченных «насыщен-постью» соответствующих биогеоценозов и эдафических пространств.
Сказанное выше можно проиллюстрировать несколькими примерами. На рис. 26 хорошо видна корреляция между числен-ностями жертвы (заяц) и хищника (рысь, лисица, волк); зависимость осенней численности землероек от метеорологических условий предыдущей весны и зависимость численности белки от урожая семян ели в предыдущем году, а также изменения численности горностая в разные годы в разных районах Якутии (последний пример интересен тем, что показывает возможность независимого колебания численности в соседних популяциях).
На рис. 27 в схематизированной форме приведено распределение во времени годов с «мышиной напастью» в России более чем за сто лет, с 1820 по 1930 г. На рис. 28 изображены крупномасштабные (1 : 1800) изменения численности бабочки Dendrolimus и, наконец, на рис. 29 — сезонные колебания численности большой синицы (Parus major); на последнем рисунке видно, что, несмотря на колебания численности по сезонам, весенняя численность остается год от года примерно одинаковой. В завершение этой серии примеров на рис. 30 в эксперименте показаны формы типичных кривых изменения численности в пределах одной «волны жизни» в системе «хищник — жертва».
В некоторых случаях удается наблюдать и описать в природе расширение или сокращение ареалов некоторых видов в очень больших масштабах; в этих случаях «волны жизни» популяций (обычно на одной из границ ареала) являются своего рода «пусковыми механизмами» и сопровождающими экспансию факторами. На рис. 31 приведено несколько примеров таких территориальных экспансий различных видов, связанных с волнами жизни. Приведенные примеры показывают быстрые вспышки расселения, неизбежно связанные со столь же резким увеличением численности иноземных видов, попадающих в условия, благоприятствующие их размножению и расселению главным образом из-за отсутствия естественных врагов.
Следующие примеры иллюстрируют быстрое расселение и связанное с этим повышение численности в естественных условиях, выражающиеся в расширении в определенном направлении видового ареала. На рис. 32 приведено расселение на северо-восток канареечного вьюрка (Serinus canaria) с 1800 до 1925 г. из его исходного средиземноморского ареала до юго-восточных побережий Балтийского и северо-западных Черного моря; европейскими орнитологами расселение канареечного вьюрка за этот период точно прослеживалось от десятилетия к десятилетию (Майр, 1926). В дальнейшем вплоть до настоящего времени канареечный вьюрок продолжал распространяться все дальше на восток и достиг сейчас нижнего и среднего Днепра. Распространение на север и восток за последние 130 лет зайца-русака (Lepus europacens), по-видимому, можно считать случаем проникновения этого вида на создаваемые человеком культурные ландшафты.
Для изученных животных и растений можно привести большое количество таких примеров; достаточно упомянуть более или менее количественно изученные расширения ареалов на север у вороны, галки, сороки и домового воробья в пределах Центра Европейской части СССР (Смолин, 1948). На рис. 33 приведен своеобразный пример волн жизни, показывающий распределение и величину очагов массового размножения водяной крысы (Arvicola terrestris) в лесостепной полосе Западной Сибири на протяжении семи смежных лет.
Еще С. С. Четвериков указал на возможное эволюционное значение «волн жизни», часто связанных с соответствующими флуктуациями занимаемых популяциями территорий; иногда они сопровождают относительно быстрые расширения (или сокращения) ареалов видов или их значительных частей. С. С. Четвериков основное эволюционное значение «волн жизни» видел в том, что этот флуктуирующий (и в этом смысле случайный) фактор влияет на направление и интенсивность давления отбора; как мы увидим далее, еще больше его значение как фактора, влияющего на случайные колебания концентраций разных генотипов и мутаций в природных популяциях.
Особенно интересны волны жизни в маргинальных популяциях, на границах видовых ареалов; они, несомненно, являются, с одной стороны, «передовыми отрядами» во взаимоотношениях вида с новыми для него комплексами биогеоценозов и физико-географических условий, а с другой стороны, как было показано Н. И. Вавиловым на огромном материале ареалов культурных растений, они являются местами выделения популяций, в высокой степени гомозиготных по некоторым рецессивным признакам, встречающимся в основной части видовых ареалов лишь в гетерозиготном состоянии и в относительно невысоких концентрациях. Таким образом, популяционные волны на границах видовых ареалов могут являться своеобразной «апробацией» новых генотипов. На рис. 34 в схематизированном виде приведены колебания с образованием и исчезновением популяционных «островков» на границе вида; в основу положены реальные данные.
Таким образом, популяционные волны, встречающиеся во всех популяциях любых живых организмов, имеют троякое значение.
Прежде всего волны жизни совершенно случайно, статистично и резко изменяют концентрацию всех редко встречающихся в популяциях мутаций и генотипов. Действительно, восстанавливающаяся после спада численности популяция включит в свой состав лишь те мутации и генотипы, которые в определенных количественных отношениях присутствуют в репродуктивной совокупности, из которой вновь нарастает численность популяции. При этом, естественно, ряд присутствовавших в малых концентрациях мутаций совершенно случайно (безотносительно к их селективной ценности) исчезнет из популяции, а другие, также находившиеся в предыдущем пике в очень малых концентрациях и случайно оставшиеся в значительно более высокой концентрации ко времени спада, резко повысят свою концентрацию. Популяционные волны, следовательно, совершенно случайно, но резко меняют набор мутаций и особенно их концентрацию в популяциях; часть мутаций случайно исчезает из популяции, а некоторые могут, также случайно, резко повышать концентрацию. Схематически это изображено на рис. 35. Случайное резкое повышение концентраций редких в популяции мутаций имеет очень существенное значение. Как мы увидим при рассмотрении естественного отбора, отбор (как положительный, так и отрицательный) очень медленно изменяет весьма низкие (или весьма высокие) концентрации мутаций и относительно быстро. Изменение происходит при исходном действии на средние концентрации (т. е. на концентрации между пятью и девяносто пятью процентами). В этом смысле популяционные волны являются поставщиком эволюционного материала под действие относительно интенсивного отбора.
|
|
abcdef abcdef abcdef abcdef abcdef abcdef aBcdef abcdEf abcdef abcdef abcdef abcdef abcdef abcdef abcdef |
aBcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef aBcdef abcdef |
abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdeF abcdef abcdef abcdef abcdef abcdef abcdef abcdEf |
abcdef abcdef aBcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef |
abcdef Abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdef abcdEf abcdef abcdef abcdef abcdcf abcdef |
abcdef abcdef abcdef abcdef abcDef abcdef abcdef abcdef abcdef abcdef abedеf Abcdef abcdef abcdef abcdef |
Рис. 35. Схема возникновения случайных изменении концентраций мутаций в популяции при действии популяциопных волн.
а, Ь, с, d, е, f — различные гены, в совокупности обозначающие индивидуум; А, В, С, D, К, F — соответствующие мутации, встречающиеся в популяции в незначительной концентрации. Если в результате волн численности сохранятся только выделенные wdtnjv группы особей (составляющие около 10% всей совокупности), концентрация мутаций в популяции резю изменится (из Бауэра, Тимофеева-Ресовского, 1943)
Во-вторых, популяционные волны характеризуются на восходящей части кривой численности во времени понижением, а на нисходящей части — повышением относительной смертности потомства из процессов повышения и понижения численности в чреде поколений. Это обстоятельство должно приводить к сопровождающему популяционные волны колебанию давлений естественного отбора (снижению на восходящей и повышению на нисходящей части кривой). Супругами Форд в этой связи очень четко было показано увеличение разнообразия и концентраций аберраций у бабочки Melitea на восходящих частях и их резкое исчезновение на нисходящих частях популяционных волн (Ford, Ford, 1930). В этом смысле популяционные волны опять-таки являются фактором, подставляющим редкие мутации под действие отбора.
И, наконец, в-третьих, популяционные волны в тех случаях, когда они сопровождаются флуктуациями, а иногда и резким расширением популяционных ареалов, выводят, хотя и временно, ряд мутаций и генотипов в другую констелляцию абиотической и биотической среды; это также является своего рода «апробацией» для ряда генотипов, способствуя «выходу» некоторых из них на эволюционную арену.
Популяционные волны, действуя совершенно иначе, чем мутационпый процесс, являются, однако, вместе с ним фактором — поставщиком эволюционного материала, выводящим ряд генотипов, совершенно случайно и ненаправленно, в качестве «кандидатов» на роль новых звеньев в протекающих эволюционных явлениях и процессах, обогащающих основной генофонд населения вида.
Давление этого фактора может быть весьма различно даже в разных популяциях одного и того же вида и, вероятно, превышает таковое спонтанного мутационного процесса.