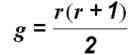
Солбриг О., Солбриг Д.
Москва, "Мир", 1982
|
|
Рекомбинацией называют смешение у потомков генов и хромосом их родителей. Строго говоря, рекомбинация не относится к числу факторов, изменяющих частоту генов в популяции. Однако она имеет важное значение главным образом по двум причинам, Во-первых, разные гены растения или животного взаимодействуют между собой, и некоторые сочетания генов оказываются при этом лучше, чем другие. Во-вторых, число рекомбинантов неизмеримо выше возможного числа мутаций. Хотя в конечном счете источником новой генетической изменчивости служат мутации, возникновение в популяции новых типов по большей части бывает обусловлено рекомбинацией. Этот процесс может создавать большее число более разнообразных форм и с большей скоростью, чем мутационный процесс, и непосредственным источником изменчивости в популяции служат рекомбинации, а не мутации. Раздельнополость представляет собой внешнее выражение модификаций, происшедших в фенотипе, для того чтобы обеспечить рекомбинацию. Рекомбинация имеет место в тех случаях, когда ядра и хромосомы мужской и женской гамет сливаются, образуя ядро зиготы.
Если гетерозиготный организм, имеющий генетическую конституцию A1A2B1B2, скрестить с другим организмом, несущим такое же сочетание генов, то их потомки (при отсутствии сцепления между двумя локусами) будут иметь (в соответствии со вторым законом Менделя) следующие генотипы: A1A1B1B1; A1A1B1B2; A1A1B2B2; A1A2B1B1; A1A2B1B2; A1A2B2B2; A2A2B1B1; A2A2B1B2; A2A2B2B2. Всего девять разных типов. Каждый из этих типов может возникнуть в результате мутации, но вероятность возникновения всех девяти типов путем мутаций в одном поколении очень мала — так мала, что это практически невозможно. (Если принять частоту мутаций равной 10-24, то вероятность возникновения этих девяти типов в популяции, состоящей из особей A1A2B1B2, за одно поколение в результате мутаций равна I0-24N, где N — число особей в данной популяции.)
Число диплоидных генотипов, которые можно скомпоновать из любого числа аллелей какого-либо гена, описывается выражением
где g — число диплоидных генотипов, а r — число аллелей в каждом локусе.
Если рассматривать два локуса, расположенных в разных хромосомах так, что они могут свободно рекомбинировать, общее число возможных диплоидных генотипов с участием этих двух генов составит
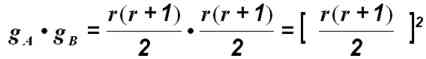
Если, например, принять, что r = 2, т. е. в каждом из двух локусов имеется по два аллеля, то
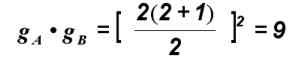
что, как мы видели, соответствует числу возможных сочетаний аллелей при рекомбинации у двух организмов, гетерозиготных по двум локусам, в каждом из которых имеется по два аллеля.
Если вместо двух рассматривать три гена, то число рекомбинаций будет равно произведению gA?gB?gC и т. д.; таким образом, для n генов (локализованных в разных хромосомах) число рекомбинантов составит
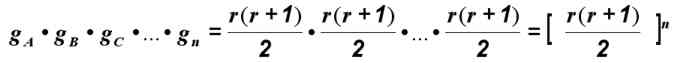
Число возможных рекомбинаций зависит, следовательно, от числа аллелей в данном локусе (r) и от числа независимых локусов (n); оно довольно велико, если r и n больше трех, как это часто бывает. Например, при n = 10 и r = 3 число рекомбинаций равно 610 = 60 466 176. Ясно, что изменчивость, получающаяся за счет рекомбинации, очень велика, так велика, что в процессе эволюции возник целый ряд механизмов для регуляции и ограничения рекомбинации.
Интересным примером, демонстрирующим важную роль рекомбинации, служит гриб, вызывающий ржавчину пшеницы (Pucciniagraminis). Диплоидные (дикариотические, т. е. содержащие по два гаплоидных ядра на клетку) споры этого гриба заражают пшеницу (а также овес, рожь, ячмень и многие другие злаки) весной. Внешние признаки заражения хорошо заметны: это вытянутые в вертикальном направлении красновато-коричневые или черноватые зернистые образования на стебле и листьях, по которым заболевание и получило название «ржавчины». На растениях пшеницы гриб продуцирует споры двух типов. Споры одного типа, называемые уредоспорами, диплоидные, они продуцируются в течение всего лета и могут заражать другие растения пшеницы. К концу лета гриб производит вместо уредоспор споры другого типа — телейтоспоры, Каждая телейтоспора претерпевает мейоз и производит четыре гаплоидные споры, называемые базидиоспорами. Мейоз и образование базидиоспор нередко происходят только следующей весной, а до тех пор телейтоспоры продолжают оставаться на отмерших растениях пшеницы или падают на землю. Базидиоспора неспособна заражать пшеницу и может дать начало мицелию (вегетативное тело гриба, грибница) лишь в том случае, если она попадет на растение барбариса. Оказавшись на листе или побеге барбариса, базидиоспора развивается в гаплоидный мицелий. В конце концов два гаплоидных мицелия разного пола (или разного типа скрещи_ ваемости, как это обычно называется у грибов) объединяются, образуя диплоидный мицелий, все еще на барбарисе. А затем этот диплоидный мицелий дает начало диплоидным спорам — так называемым эцидиоспорам, заражающим растения пшеницы (они не способны заражать барбарис), и весь цикл возобновляется (рис. 6.5).
 |
|
Рис. 6.5. Жизненный цикл ржавчины (Puccinia graminis). |
Споры (эцидиоспоры) прорастают на пшенице, образуя мицелий, состоящий из двуядерных клеток. Этот мицелий дает начало уредоспорам, которые могут заражать другие растения пшеницы. В конце сезона продуцируются телеитоспоры. Два содержащихся в тедейтоспоре ядра сливаются и в таком состоянии гриб перезимовывает. Весной телеитоспоры претерпевают мейоз, в результате чего образуются гаплоидные базидиоспоры, которые способны прорастать только на барбарисе. Из базидиоспор развивается гаплоидный мицелий, дающий начало спермогониям и рецептивным гифам. Клетки из спермогониев переносятся на эти гифы, где образуются двуядерные эцидиоспоры, которые могут прорастать только на пшенице.
Переход с одного вида хозяина на другой в жизненном цикле ржавчины пшеницы имеет интересные эволюционные и экономические последствия. Рекомбинация не может происходить в отсутствие второго растения-хозяина — барбариса. Кроме того, в областях с суровой зимой все уредоспоры погибают от холода, и без растений барбариса ржавчина не может выжить. Поэтому в таких областях (например, на пшеничных полях Дакоты и центральных провинций Канады) искоренение кустов барбариса задерживает весной заражение пшеницы ржавчиной; Эта мера не может совершенно ликвидировать заражение, потому что уредоспоры приносятся ветром из областей с более мягким климатом, где перезимовавшие уредоспоры уже успели заразить растения пшеницы и начали продуцировать новые споры. Очень важно, однако, что искоренение барбариса элиминирует рекомбинацию, происходящую при половом процессе у ржавчины. Современные сорта пшеницы иммунны, или высоко устойчивы, к заражению ржавчиной. Но в процессе эволюции непрерывно возникают новые генотипы ржавчины, устойчивостью к которым пшеница не обладает, что заставляет заниматься созданием все более устойчивых сортов. Генетическая изменчивость в популяции ржавчины возникает в результате мутаций и рекомбинации. Устранение возможности половой рекомбинации, достигаемое путем искоренения барбариса, замедлит процесс эволюционного приспособления у ржавчины. Этот пример немногим отличается от ситуации с вирусом гриппа у Homo sapiens. Мы в состоянии справиться с вирусом, который живет в популяции человека и может преодолеть его защитные механизмы только при помощи мутации; однако вирусы, возникающие в результате рекомбинации вирусов человека с вирусами других видов, например обезьян или свиньи, представляют серьезную проблему.